Originea haplogrupului I
Originile vechilor europeni pot fi, fără îndoială, identificate cu ADN haplogrupul I, separat de Y care a venit din Orientul Mijlociu prin haplogrupul IJ și C6, care erau Cro-Magnons.În genetica populației umane, (M170, P19, M258) Y este un haplogrup cromozomial care se presupune că a apărut acum 20-25 de mii de ani în Peninsula Balcanică.

apare la aproape 1/5 din populația europeană. Unii cercetători cred că locul de origine haplogrupul I s-ar putea să fi existat o cultură Gravettiană, un fel de „oază” locuită în care oamenii au experimentat Ultimul Maxim glaciar, care a avut loc acum aproximativ 21 de mii de ani. De la sosirea oamenilor moderni (Cro-Magnons) în Europa cu aproximativ 40 de mii de ani în urmă și până la invazia indo-europeană de acum aproximativ 5 mii de ani, se pare că, haplogrupul I a constituit majoritatea populației europene, s-ar putea să fi existat un număr mai mare de subclade, acum dispărute, dintre care doar două au fost identificate acum (I1 și I2). Constructorii de megaliți din Europa de Vest au aparținut cel mai probabil.
haplogrupul I2a
Distribuția etnogeografică a haplogrupului I Unii cercetători cred că locul de origine Cel mai răspândit există în 2 focare de distribuţie: primul este haplogrupul I1centrat în țările scandinave printre suedezi, finlandezi, danezi și norvegieni. Al doilea este haplogrupul I2
centrat pe Balcani din Croaţia şi Serbia. există în 2 focare de distribuţie: primul esteŞI este un haplogrup european indigen. Majoritatea purtătorilor moderni ai haplogrupului I1 sunt vorbitori de limbi germanice ale familiei indo-europene, deși inițial acest haplogrup a fost asociat nu cu popoarele indo-europene, ci cu substratul pre-germanic (posibil Cultura Funnel Beaker) . I1 este identificat de cel puțin 15 mutații unice, ceea ce înseamnă că acest grup fie a fost complet izolat pentru o perioadă lungă (ceea ce este puțin probabil) sau a experimentat un blocaj serios în vremuri relativ recente. Deși prima mutație care s-a separat I1 de la mine, ar fi putut să apară acum 20 de mii de ani, toți purtătorii de astăzi ai acestui ADN haplogrupuri provin de la un om care a trăit nu mai devreme de 5 mii de ani în urmă. Acest lucru coincide bine cu sosirea indo-europenilor în Scandinavia, despre care se crede că au ucis cea mai mare parte a populației masculine indigene sau au pus familiile lor într-un dezavantaj demografic. Așadar, pare destul de plauzibil că o singură familie de indigeni scandinavi a supraviețuit acestei invazii (sau, de exemplu, poate un băiat), ai cărui descendenți au alcătuit mai târziu haplogrupul I1, care a devenit astfel o marcă de încredere a Peninsulei Scandinave și a celor care s-au format pe formațiunile etnice din acea epocă. Astăzi, reprezentanții acestui grup se găsesc aproape peste tot în Europa, unele subclade răspândite cu migrațiile germane, singura excepție poate fi nordul părții europene a Rusiei (Karelia, Vologda) și cea mai mare parte a regiunii Volga, unde a fost transmis haplogrupul I1. prin finlandezi. Haplogrupul I2 are 3 subclade principale cu distribuții etnogeografice diferite în Europa:
Subcladele I2-M26 este una dintre principalele linii cromozomiale Y din Sardinia (atinge 40%) și printre basci. De asemenea, se găsește cu frecvență scăzută în toată Europa de sud-vest: Spania, Franța, Portugalia, Italia.
Subclada I2-M423 este caracteristică populațiilor din Balcani și Carpați, cel mai des observată în rândul slavilor dinarici (sârbi, croați și bosniaci), precum și în Moldova și sud-vestul Ucrainei. De asemenea, se găsește cu o frecvență notabilă în Albania, nordul Greciei, Bulgaria, Slovacia, estul Ucrainei, Belarus și sud-vestul Rusiei.
Subclada I2-M223 este caracteristică nord-vestului Europei și atinge un maxim în Germania (Saxonia Inferioară). Intalnit si in Romania, Moldova si cu frecventa redusa in Europa de Est.
Haplogrupuri de europeni moderni după expansiunea indo-europeană și Marea Migrație

Printre etnicii europeni moderni, următoarele haplogrupuri sunt comune în ordine descrescătoare (inclusiv sudul și estul Europei): haplogrup R1b1a2 (33,71%), haplogrup R1a1 (22,21%), există în 2 focare de distribuţie: primul este(8,39%), haplogrup I2a (7,97%), haplogrup E1b1b1 (6,77%), haplogrup J2 (6,48%), haplogrup N1c1 (5,82%), haplogrup G2a3 (3,92%), haplogrup I2b (2,20%) și cu frecvență scăzută se găsesc haplogrupurile T (1,82%), Q1a (1,32%) și J1 (0,85%). Haplogrupurile cu o frecvență totală mai mică de 0,5% nu sunt prezentate în diagramă.
Aceste procente de haplogrupuri în rândul europenilor iau în considerare dimensiunea populației indigene a țărilor europene cu o populație de peste 1 milion de oameni și frecvența medie a haplogrupurilor pentru acestea din tabelul de mai jos, eroarea totală nedepășind 5%.

Aceste haplogrupuri cu frecvențe variabile de la 0-95% se găsesc printre germani, spanioli, italieni, greci, polonezi, maghiari, francezi, albanezi, englezi, irlandezi, scoțieni, portughezi, elvețieni, cehi, slovaci, sârbi, croați, români, belaruși. , ucraineni, danezi, suedezi, norvegieni, finlandezi, ruși, estonieni, letoni, lituanieni etc.
genetica europeana
Haplogrupurile în sine nu poartă informații genetice, deoarece Informația genetică este localizată în autozomi - primele 22 de perechi de cromozomi. Vedeți distribuțiaOriginea haplogrupului cromozomial Y I2a, evoluția sa și rutele de migrație ale populațiilor de purtători ai acestui haplogrup au făcut obiectul studiului meu special timp de 5 ani, începând din momentul în care am devenit interesat de studiul activ al populației umane. genetica.
Motivul interesului meu pentru acest haplogrup este destul de simplu - o analiză genetică a cromozomului meu Y (Y-STR + Y-snips) a arătat că, din punct de vedere al clasificării filogenetice, acesta aparține uneia dintre ramurile acestui haplogrup și datorită acestui fapt biologic banal, în ea descind din linia masculină directă de la reprezentanții antici ai acestui haplogrup. Un motiv mai puțin important pentru implicarea mea în studiul acestui haplogrup este nefericita necunoaștere atât a I2, cât și a întregului haplogrup I. E suficient să spunem că cele mai recente studii speciale asupra acestui haplogrup au fost publicate în 2004, 2006, 2007 - Rootsi , Siiri et al.(2004), Rootsi, S. (2006), Peter Underhill et al (2007). Lucrările rămase, în special cele referitoare la diversitatea cromozomială Y a populațiilor din Balcani și din fosta Iugoslavie, repetă în principal concluziile exprimate în cele trei lucrări de mai sus, fără a adăuga nimic propriu (Marjanović, Damir; et al.2005, Peričić). , Marijana et al. 2005; Rebala K., et al.(2007). Prin urmare, munca cercetătorului popgeneticist eston Rootsi a fost cea care a determinat o serie de ipoteze de bază privind originea haplogrupului I, care au fost ulterior consolidate în sursele științifice și populare (Wikipedia). Din păcate, multe dintre aceste ipoteze s-au dovedit a fi false.
Mai jos vom prezenta câteva prevederi cuprinse în articolul Wikipedia despre I2 care sunt în general acceptate în genetica oficială a populației conservatoare și, judecând după articol, unele dintre prevederile de mai sus au fost ulterior regândite și dezvoltate în cercurile apropiate de genetică ale amatorilor.
Haplogrupul I2 atinge cea mai mare concentrare în rândul locuitorilor din Bosnia, Serbia și Croația - acum aceste popoare vorbesc limbi slave, totuși, concentrarea haplogrupului „indo-european” R1a printre ei este minimă, ceea ce ne permite să vedem în ele direct descendenții substratului preslav, care au fost ilirii și tracii, precum și strămoșii lor, sunt purtători ai culturii neolitice balcanice (inclusiv cultura Tripoli). Ultima teză este confirmată de faptul că atât neoliticul balcanic este o dezvoltare a Orientului Mijlociu, cât și haplogrupul I balcanic este înrudit cu haplogrupul J din Orientul Mijlociu, deși datele despre divergența lor variază din Epoca Bronzului (Cultura câmpurilor de urne funerare: Peștera Liechtenstein), care, totuși, a zdruncinat oarecum ideile tradiționale că la acea vreme Europa era locuită de celți - purtători fie ai haplogrupului „celtic” R1b, fie ai „arienilor”. R1a. Relația dintre haplogrupul balcanic I2 și haplogrupul scandinav I1, cu populațiile lor fragmentate, ne permite să concluzionăm că au fost disecate prin invazia popoarelor haplogrupurilor „celtice” (sau mai degrabă iberice).
Lăsând deoparte problema conexiunii dintre haplogrup și limbă, haplogrup și cultura arheologică (aceste legături merită un studiu separat), vom vedea că principalul argument al lui Rootsi în favoarea patriei ancestrale balcanice I este prezența semnificativă a acestui haplogrup în limba dinarică. Regiunea Alpi/Balcani. Cea mai simplă și mai ieftină, din punct de vedere al parcimoniei, explicație pentru astfel de numere este asumarea continuității unei părți a fondului genetic al populației balcanice de la momentul ultimului maxim glaciar din Europa (acum 20 de mii de ani), adică, încă de pe vremea refugiului glaciar balcanic. După cum se știe, originea geografică a haplogrupurilor de cromozomi Y poate fi stabilită prin următorul criteriu empiric: în locul sau populația în care a apărut un anumit haplogrup, acesta frecvența și dispersia STR (sau vârsta variabilității STR) maximă comparativ cu alte populații (Sengupta și colab., 2006). Această schemă simplă și aparent logică, atunci când este examinată mai detaliat folosind exemplul haplogrupului I, nu rezistă criticilor în lumina noilor date. Merită să ne amintim, totuși, că contextul din articolele lui Rootsi a fost stabilit într-un moment în care geneticienii aveau doar o înțelegere generală a structurii haplogrupului I. Cercetări ulterioare efectuate de geneticieni specializați ai populației, cuplate cu studiul amator dedicat al haplogrupului I în proiecte comerciale FTDNA (Family Tree DNA - Haplogroup I Y-DNA Project, FTDNA I1 yDNA Haplogroup, FTDNA I2*New ISOGG I2b, FTDNA I2a Y-Haplogroup, FTDNA I-M223 Y-Haplogroup, FTDNA I2a2b-L38) și eforturi deosebit de semnificative de cercetători amatori individuali în genealogie numită ADN (Kenneth Nordtvedt, Bernie Cullen, Steve Trangsrud, William Hartley, Yme Drost, Aaron J. Hill, Peter Hrechdakian, Hovann Simonian, Bob May, Vadim Verenich, Bill Morrow, Lawrence Mayka, Vladimir Semargl etc.) a permis o mai bună și a obține o înțelegere mai profundă a relațiilor filogenetice și a structurilor ramurilor-subcladelor haplogrupului, precum și a oferi o referință cronologică și geografică pentru ramurile-subcladele individuale ale haplogrupului. Acest studiu se va limita doar la un scurt instantaneu al stării actuale a cunoștințelor noastre despre originea și „evoluția” haplogrupului I2a.
Originea și rutele de migrare ale haplogrupului I2a.
Datorită naturii fragmentare a revizuirii, nu vom atinge întrebări la fel de importante despre cum și când haplogrupul I a apărut în Europa, precum și întrebări despre momentul și locul divizării ramului bazal I în haplogrupurile I1 și I2. În prezent, nu avem suficiente dovezi în favoarea uneia dintre numeroasele și la fel de probabile versiuni ale scenariului și, prin urmare, pur și simplu vom ocoli aceste întrebări în speranța unui viitor studiu al ADN-ului locuitorilor din Europa paleoliticului mijlociu.
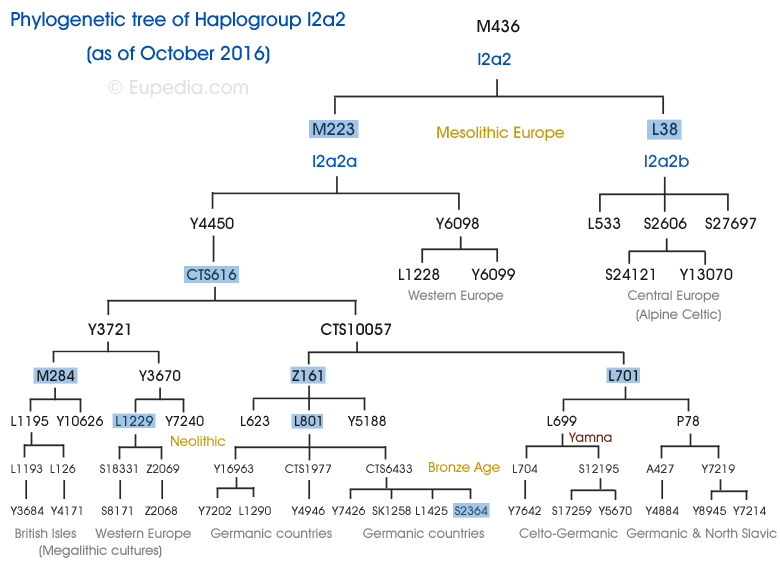
În schimb, vom începe descrierea prezentând arborele filogenetic („arborele genealogic”) al haplogrupului I2a, lăsând deoparte celelalte ramuri ale acestui grup. În loc de diagrame simple standard ale haplogrupului I, oferim o versiune canonică a arborelui în procesarea lui Nordtvedt (versiunea 2013). Principalul său avantaj este reprezentarea vizuală a aranjamentului topologic relativ al ramurilor subcladelor și al clusterelor intraclade de haplotipuri. În plus, scara de sub arbore este construită ținând cont de timpul intercladei de divergență (divergență) a ramurilor, calculat folosind metoda dispersiei încrucișate de Ken Nordtvedt. În fine, reprezentarea grafică a arborelui filogenetic este completată de o reconstrucție conjuncturală a mișcării purtătorilor haplogrupului I în cronotopul Europei preistorice.
Cronologia diviziunii haplogrupului I2a este următoarea (versiunea noastră):
Acum aproximativ 14.000 -15.000 de ani din I2a L460/PF3647/S2381 a apărut o ramură I2a2-L35/PF3862/S150, L37/PF6900/S153, L181, M436/P214/PF3856/S33, P216/PF3855/S30, P217/PF3854/S23, P218/S32, care aproape imediat s-a împărțit în I2a2a-L34/PF3857/S151, L36/S152, L59, L368, L622, M223, P219/PF3859/S24,
P220/S119, P221/PF3858/S120, P222/PF3861/U250/S118, P223/PF3860/S117, Z77(aprox. 12.000 - 8.000 de ani) și I2a2b L38/S154, L39/S155, L40/S156, L65.1/S159.1, L272.3(aprox. 12.000 -8.000 de ani).
În paralel cu aceasta, a apărut o subramură din haplogrupul I2a I2a1-P37.2 (15.000-10.000 de ani înainte de prezent). Este de remarcat faptul că în timpul analizei extinse a fost identificat un haplogrup separat I2a1 -F (snips de identificare L1294 L1297 L1298 nu sunt incluse în arborele ISOGG). Conform vârstelor interclade calculate de Nordtvedt, acest cluster a apărut mai târziu decât grupul I2a1a, dar mai devreme decât grupul I2a1b (adică, la mijlocul dintre cele două ramuri). Potrivit lui Nordtvedt, ultimul strămoș comun al lui I2a1-F și I2a1a ar fi putut trăi nu mai târziu de 20.000 de ani înainte de vremea noastră. Este caracteristic că reprezentanții I2a1*-F trăiesc în principal pe teritoriul vestului modern al Franței. În plus, reprezentanții I2a1c locuiesc și în Europa de Vest.
Din I2a1-P37.2între 20.000 și 15.000 î.Hr cutie timpul lui s-a remarcatI2a1aL158/PF4073/S433, L159.1/S169.1, M26/PF4056((durata de viață a ultimului strămoș comun - - 8.000-6.000) șiI2a1b-M423 L178/S328, M423(durata de viață a ultimului strămoș comun este cu nu mai puțin de 14.000 de ani înainte de prezent), de la I2a1b-M423 L178/S328 conform relatării interclade, liniile ancestrale au fost distinse cu aproximativ 13.500 de ani în urmăI2a1b3 și I2a1b2 - (Dinaric+Disles) și Insulele. În mod interesant, estimăm că durata de viață a ultimului strămoș comun al celui dintâi este cu aproximativ 6600 de ani înainte de momentul prezent pentru Insule la aproximativ 8000 de ani.

Mai mult decât diviziuni intraclade târzii (atexact 8 mii-6 mii de ani înainte de prezent) se corelează bine cu răcirea globală a cca. 6200 î.Hr., care a dus la criza demografică neolitică remarcată de experți, când populația din Europa a scăzut în mod clar.În nordul Europei în același timp vremea mea (aproximativ 6500 -6200 î.Hr.) a avut loc un eveniment care a separat geografic (separate filogenetic mai devreme) populațiile I2a1b2-Isles și I2a1b3 , precum și o parte a subcladei I2a2-M284 – Isles din subcladele I2a2 rămase.Vorbim despre inundațiile din Doggerland. Definit cudovezile, inclusiv contururile fundului mării moderne, arată că, în timpul ultimei glaciații, distribuția apelor dintre Marea Nordului și Canalul Mânecii se întindea de la East Anglia la sud-est până la Hoek van Holland, mai degrabă decât prin Pas de Calais, și că râurile Tamisa, Meuse, Scheldt și Rinul, unite, curgeau de-a lungul Canalului Mânecii sub forma unui râu larg, care în cele din urmă se varsă în Oceanul Atlantic.[ În jurul anului 8000 î.Hr. e. coasta de nord a masei uscate, Doggerland, era o coastă cu lagune, mlaștini, țărmuri de maree și plaje. Poate că în perioada mezolitică această zonă a fost cea mai bogată din Europa în ceea ce privește vânătoarea, păsările și pescuitul.
Se presupune că o creștere treptată a nivelului mării ca urmare a topirii ghețarului (sfârșitul ultimei glaciații) a dus la inundarea Doggerland de către Marea Nordului, care a separat treptat Marea Britanie preistorică de continentul european în jurul anului 6500. î.Hr. e. Pe locul viitorului banc de nisip, cunoscut în prezent sub numele de Dogger Bank, conform acestei ipoteze, insula a continuat să existe până cel puțin în anul 5000 î.Hr. e.
O ipoteză alternativă, recent propusă este că cea mai mare parte a terenului a fost inundată de un tsunami în urmă cu aproximativ 8.200 de ani (6.200 î.Hr.) cauzat de unul una dintre cele mai mari din istorie alunecări de teren - deriva zonei de sol subacvatic de la coasta Norvegia cunoscută sub numele de Sturegga . Conform acestei teorii, tsunami-ul provocat de alunecarea de teren a jucat un rol catastrofal pentru populația mezolitică a zonei. Ca urmare a tsunami-ului, Marea Britanie pare să fi fost separată definitiv de continent și, din punct de vedere cultural, mezolitic a mers pe drumul lui pe el. Unul dintre efectele secundarea avut loc o alunecare de teren răcire globală 6200 î.Hr e. datorita afluxului de apa rece din ghetarii topiti.
Cine au fost reprezentanții preistorici ai haplogrupului I2a?
eu. Unde au apărut strămoșii haplogrupului I2a??
După cum se poate observa din citatul de mai sus de pe Wikipedia, printre unele cercuri genetice, opinia incorectă în opinia noastră că I2a este asociată cu cercul culturilor neolitice balcanice, inclusiv culturi apropiate de Trypillians, a devenit mai puternică. Această opțiune este posibilă, dar versiunea finală ar trebui determinată prin analize ale cromozomilor Y mai vechi decât ADN-ul rămășițelor reprezentanților acestor culturi. Dar, dacă I2 au fost prezenți în rândul tripilienilor, ei cu greu pot fi considerați un element indigen (al cărui miez erau imigranții din Orientul Mijlociu, ei reprezentau rămășițele unui element mezolitic local încorporat în această cultură); Dacă vorbim despre cultura primordială a anticului I2a2, atunci aceștia erau cel mai probabil sviderienii și epigonii lor.
În 2011, am putut avea o mică discuție cu administratorul proiectului polonez FTDNA (Larry Majka) despre originea haplogrupului I2a1b-M423 (și I2a în general). Înainte de aceasta, Mike era relativ puțin interesat de originile acestui haplogrup (deși, la fel ca mine, el aparține acestuia). El a propus o versiune extrem de interesantă despre locul de origine al haplogrupului I2a1. În opinia sa, strămoșul comun al tuturor I2a1 (ca I2a1*, I2a1a, I2a1b, I2a1c) cu un grad ridicat de probabilitate ar fi putut trăi în regiunea alpină.
De asemenea, a fost de acord cu traseul propus de mine pentru trecerea clanului I2a1 din Alpi în Carpați. Adevărat, nu a fost de acord cu întâlnirea mea. Cred că populația ancestrală din I2a1 și-a început mișcarea din Alpi spre Carpați (din Europa de Vest spre nord în urma ghețarului în retragere), imediat după sfârșitul LGM (adică în mezolitic undeva după 10.000 î.Hr.), Mike credea că că inițiatorul migrației ar fi putut fi celții (adică această migrație s-a produs deja în epoca bronzului). Ultima opțiune, deși interesantă, nu explică momentul și locul separării subcladei insulare I2a1b2 și subcladei dinarice I2a1b3, care, conform datării moleculare, nu ar fi putut avea loc mai târziu de 10-8 mii de ani în urmă.
Pentru a verifica versiunea lui Mikey, am decis să folosesc algoritmul Mesquite, care ne permite să estimăm locul cel mai probabil (din punct de vedere al parcimoniei) al „fundației” subcladei, pe baza distribuției geografice actuale (geografice). răspândire) a două subclade adiacente (în acest caz, determinăm locul apariției I2a1b1 pe baza unei comparații a coordonatelor geografice ale taxonilor haplotip și topologia arborelui I2a1b, care a fost înrădăcinată de rădăcina I2a1a exogrupului).
În ciuda faptului că, în prezent, cea mai mare densitate a subcladei I2a1a este în Sardinia, conform opiniei bine motivate și confirmate empiric a lui Nordtvedt, cele mai vechi clustere I2a1a în vârstă se află în regiunea de nord. Peninsula Iberică, Alpi și părți din sudul Franței.
Prin urmare, coordonatele centrului geografic al Munților Pirinei au fost luate drept coordonatele rădăcinii exogrupului (haplotipul I2a1a)
În acest scop, am învățat cum să aplic arborii filogenetici din Mesquite direct la proiecția geografică a Europei de Nord, în proiecție 2D-3D.
De exemplu, 2 hărți pe care le-am construit în Mesquite Cartographer pe baza structurii unui arbore filogenetic de 687 de haplotipuri I2a1b-M423, identificate prin găsirea a 20 de potriviri independente dintre cei mai buni 20 de arbori (parsimonici) din TNT. Culoarea roșie indică locul de origine reconstruit - în acest caz regiunea alpină.

Astfel, traiectoria pe care am obținut-o pentru purtătorii I2a1b corespunde aproximativ cu traseul propus de noi. Este greu de spus exact când a avut loc migrația, deoarece între apariția lui I2a1b și I2a1b3 au trecut câteva mii de ani. Un alt lucru este important, am arătat că mișcarea a început nu de pe teritoriul Balcanilor, ci mai degrabă din Alpii francezi sau elvețieni spre nord-est.
Este această versiune susținută de dovezi materiale - ADN antic etc.?
Literal, până de curând, ipoteza noastră propusă a rămas doar o interpolare statistică care necesită o confirmare suplimentară.
În urmă cu câteva săptămâni, a fost publicată în sfârșit munca geneticienilor „profesioniști”, care arată cum cea mai mare parte a ADN-ului eșantioanelor antice studiate până în prezent se corelează cu ADN-ul populațiilor umane moderne ( Genomul uman antic sugerează trei populații ancestrale pentru europenii de astăzi). Ipoteza mea despre punctul de plecare al originii și începutul migrațiilor lui I2a1b, pe care am exprimat-o în ultimii ani, a primit confirmare materială în lucrare.
În primul rând, în linii uniparentale, vechiul grup de gene ADN al vânătorilor-culegători din mezolitic suedezi și luxemburghezi s-a dovedit a fi reprezentanți ai haplogrupului ADNmt U și ai haplogrupului I cromozomial Y, așa că din nou avem de-a face cu absența R1a/R1b la începutul anului. Europenii mezolitici.
În al doilea rând, aceste studii arată că yuxembourgish Vânătorul-culegător mezolitic (ale cărui rămășițe au fost găsite în timpul explorării sitului mezolitic din Heffingen-Lochbourg, Luxemburg, cu aproximativ 8.000 de ani înainte de prezent) aparținea haplogrupului I2a1b* M423 + L178+. Conform datelor despre polimorfismele cromozomilor Y date în tabel, haplogrupul acestui Luxemburg preistoric este ancestral atât cu I2a1b2, cât și cu I2a1b3.
I2a1b L178 G A 15574052 12 +
I2a1b M423 G A 19096091 13 +
I2a1 P37.2 T C 14491684 7 +
I2a L460 A C 7879415 7 +
I2 M438 A G 16638804 14 +
I2 L68 C T 18700150 12 +
I P38 A C 14484379 2 +
I M170 A C 14847792 14 +
I M258 T C 15023364 5 +
I U179 G A 16354708 9 +
I L41 G A 19048602 3 +
În al treilea rând, aceste studii arată, de asemenea, că o parte din populația mezolitică din Motala (Suedia, Östgäterland, cu aproximativ 8000 de ani înainte de prezent) a aparținut și ele. I2a1b*(xI2a1b1, I2a1b3)
Haplogrupul I2 a apărut în sud-estul Europei în urmă cu peste 15 mii de ani.

Cea mai mare concentrație a sa se găsește în Bosnia-Herțegovina și în sudul Dalmației (Croația) și reprezintă, de asemenea, un procent semnificativ din toate populațiile slave de sud.
Reprezentanți ai haplogrupului I2a au stat la originile celor mai vechi culturi din Peninsula Balcanică, precum: Chel Kladove, Lepenski Vir, Gura Baciului, Nea Nicomedia. Pe baza acestui clan, se pare că s-au dezvoltat triburile iliriene și tracice. Ramura I2a a fost una dintre primele care au populat Sardinia.
Ramuri de haplogrup:
I2a1
Haplogrup I2a1(M26) reprezintă aproximativ 40% din toate descendența masculină din Sardinia. În plus, se găsește și cu frecvență scăzută până la moderată în Țara Bascilor și Peninsula Iberică și a reprezentat 1,6% (1/64) într-un eșantion de albanezi care trăiesc în Macedonia și 1,2% (3/257) într-un eșantion. Cehov Vârsta variației YSTR pentru subclada M26 este de 8,0 ± 4,0 mii de ani (Rootsi 2004).
Se presupune că ramura I2a s-a remarcat din I2 acum aproximativ 11.000 de ani în Balcani, și o ramură I2a1 s-a remarcat din I2a acum aproximativ 8.000 de ani în Sardinia sau undeva pe coasta europeană în vecinătatea ei.
I2a2

Distribuția haplogrupurilor I2a2
I2a2(snp M423) este tipic pentru populația din sud-estul Europei. Cea mai mare densitate este în Dalmația (Croația) și Bosnia-Herțegovina (>50%). Se presupune că în această zonă de pe Munţii Dinarici această ramură provine din I2a acum aproximativ 7500 de ani. Cel mai frecvent haplogrup I2a2întâlnită printre popoarele slave (în special slave de sud), și întâlnită și printre români, moldoveni, maghiari, lituanienii din sud, albanezi, greci, locuitorii din nord-estul Italiei, vestul Anatoliei și din Caucazul de Nord. Cea mai mare frecvență și diversitate de haplogrup I2a2în rândul populației din vestul Balcanilor sugerează că regiunea Adriatică a Croației moderne a servit drept refugiu pentru purtătorii acestui haplogrup în timpul Ultimul Maxim glaciar. Ramura snp P37.2 a avut loc acum aproximativ 10,7 ± 4,8 mii de ani (Rootsi 2004). Vârsta ramului snp P37.2 este de 8,0 ± 4,0 mii ani (Rootsi 2004), iar pentru snp M423 este de 8,8 ± 3,6 mii ani (Underhill 2007). Pericic consideră că răspândirea acestei ramuri a avut loc „nu mai devreme de tranziția Holocenului și nu mai târziu de Neoliticul timpuriu” (Pericic 2005). Al doilea centru de concentrare I2a2 situat în Moldova actuală și coincide aproximativ cu granițele culturii Trypillian, se pare că acest subhaplogrup era caracteristic acestei culturi, iar odată cu venirea indo-europenilor s-a alăturat fondului lor genetic, fără a experimenta o asemenea suprimare ca și alte ramuri ale haplogrupul euîn Europa centrală și de vest.
I2b1
Distribuții subhaplogrup I2b1 coincide cu distribuția haplogrupului I1, cu excepția Fennoscandia, care sugerează prezența sa în cel puțin unul dintre adăposturile paleolitice, unde a fost localizat și haplogrupul. I1. I2b1 Absența I2b1în Fennoscandia se poate datora faptului că haplogrupul I2b1 cele mai timpurii așezări din regiune au fost influențate de „efectul fondator” și de deriva genetică datorită rarității sale, deoarece haplogrupul
constituie mai puțin de 10% din diversitatea totală cromozomială Y a populației din zona specificată din afara Saxonia Inferioară. I1 Distribuția haplogrupurilor I2b1Şi I2b1 se corelează relativ bine cu granițele istorice ale distribuției vorbitorilor de limbi germanice. Haplogrup întâlnit la peste 4% din populație doar în Germania, Țările de Jos, Belgia, Danemarca, Anglia (cu excepția Țării Galilor și Cornwall), Scoția, vârful sudic al Suediei și Norvegiei, precum și în provinciile Normandia, Maine, Anjou și Perche în nord-vestul Franței, în Provence în sud-estul Franței, în regiunile istorice ale Italiei - Toscana, Umbria și Latium; precum și în Moldova, în regiunea Ryazan și în Mordovia. Se pare destul de probabil că prezența lui I1 și I2b1 în Franța, Anglia și Italia moderne, precum și în Europa de Est, este deja asociată cu expansiunea celtică și germanică, iar în vremurile preindo-europene aceste haplogrupuri erau concentrate doar în nordul Europa. Una dintre ramurile haplogrupului I2b1, și anume I1 Distribuția haplogrupurilor I2b1a(snp M284), găsit aproape exclusiv în populația britanică, ceea ce poate indica istoria sa lungă de existență în Insulele Britanice. Interesant, cu o frecvență scăzută a haplogrupului
Haplogrup I2b1 I2b
se găsesc în regiunile istorice Bitinia și Galația din Turcia modernă, unde ar fi putut fi aduse de celți, care au migrat acolo la invitația lui Nicomede I al Bitiniei. I2b1a apare și la aproximativ 1% din populația Sardiniei. I2 Se presupune că I2b1 se remarcă din I2b1aîn Europa centrală, lângă marginea unui ghețar care se retrage lent, acum aproximativ 13 mii de ani, I2b1- de la I2b1 chiar mai la nord, în ceea ce este acum Germania, acum aproximativ 9 mii de ani. Mai exact filiala britanică
s-a remarcat din
Haplogrupul I2b2 a fost descoperit în rămășițele scheletice găsite la Peștera Lichtenstein, un sit arheologic din epoca bronzului din centrul Germaniei, unde au fost găsite și artefacte din cultura Urn Fields. Dintre cele 19 rămășițe masculine din peșteră, haplogrupul I2b2 a fost găsit în 13, R1b în unul și R1a în două.
Probabil, peștera a fost situată la epicentrul răspândirii de atunci a haplogrupului I2b.
Haplogrupul I2 este cea mai comună descendență paternă în fosta Iugoslavie, România, Bulgaria și Sardinia și o descendență majoră în majoritatea țărilor slave. Frecvențele sale maxime sunt observate în Bosnia (55%, inclusiv 71% în croații bosniaci), Sardinia (39,5%), Croația (38%), Serbia (33%), Muntenegru (31%), România (28%), Moldova (24%), Macedonia (24%), Slovenia (22%), Bulgaria (22%), Belarus (18,5%), Ungaria (18%), Slovacia (17,5%), Ucraina (13,5%) și Albania ( 13,5%). Se găsește cu o frecvență de 5 până la 10% în țările germanice.
Filogeneza lui I2
Dacă sunteți nou în genealogia genetică, vă rugăm să consultați Introducerea în filogenetică pentru a înțelege cum să citiți un arbore filogenetic.
Arborele filogenetic al lui I2 a evoluat mult în ultimii 10 ani și subcladele de nivel superior au fost redenumite mai des decât pentru orice alt haplogrup, cu excepția R1b. Pentru a evita orice confuzie, se recomandă să verificați de două ori mutațiile definitorii (SNP) atunci când citiți studii mai vechi sau faceți referințe la o anumită subcladă. Mai multe subclade minore nu au fost incluse din cauza lipsei de spațiu.
- Click pentru a mari
- I2* (M438/P215/S31)
- I2a (L460)
- I2a1 (P37.2)
- I2a1a (CTS595)
- I2a1a1 (M26)
- I2a1a1a (L672)
- I2a1a1a1 (L160)
- I2a1a1a1a (PF4088)
- I2a1a1a1a1 (CTS11338)
- I2a1a1a1a1a (Z105)
- I2a1a1a1a1a1 (Z118)
- I2a1a1a1a1a1a (Z106)
- I2a1a1a1a1b (PF4189)
- I2a1a1a1a1b1 (PF4398)
- I2a1a1a1b (F1295)
- I2a1a1a1b1 (PF6950)
- I2a1a2 (L880)
- I2a1a3 (L1286)
- I2a1b (M423)
- I2a1b1 (M359.2/P41.2)
- I2a1b1a (L147.5)
- I2a1b1a1 (P41.2/M359.2)
- I2a1b2 (L161.1/S185)
- I2a1b3 (L621/S392)
- I2a1b3a (L147.2)
- I2a1c (L1286)
- I2a1c1 (L1287)
- I2a1c1a (L233/S183)
- I2a1d (L880)
- I2a1e (L1294)
- I2a1e1 (L1295)
- I2a2 (S33/M436/P214)
- I2a2a (M223)
- I2a2a1 (M284)
- I2a2a1a (L1195)
- I2a2a1a1 (L126/S165, L369)
- I2a2a1a2 (L1193)
- I2a2a2 (L701)
- I2a2a2a (P78)
- I2a2a2b (L699/L703)
- I2a2a2b1 (P704)
- I2a2a3 (Z161)
- I2a2a3a (L801/S390/Z76)
- I2a2a3a1 (P95)
- I2a2a3a2 (Z78)
- I2a2a3a2a (L1198)
- I2a2a3b (L623)
- I2a2a4 (L1229)
- I2a2a4a (Z2054)
- I2a2a4a1 (L812/S391)
- I2a2a4b (L1230)
- I2a2a5 (L1228)
- I2a2b (L38/S154)
- I2a2b1 (L533)
- I2b (L416)
I2c (L596)
Haplogrupul I este cel mai vechi haplogrup major din Europa și, după toate probabilitățile, singurul care și-a apărut acolo (în afară de haplogrupuri foarte minore, cum ar fi C1a2 și subcladele profunde ale altor haplogrupuri). Haplogrupul IJ ar fi ajuns din Orientul Mijlociu în Europa cu aproximativ 35.000 de ani în urmă, apoi s-ar fi dezvoltat în haplogrupul I la scurt timp după aceea. S-a confirmat acum că primul Homo sapiens care a colonizat Europa în perioada aurignaciană (cu 45.000 până la 28.000 de ani în urmă), a aparținut haplogrupurilor CT, C1a, C1b, F și I.
Se crede că I2 (M438/P215/S31) a apărut în timpul Paleoliticului târziu, în jurul perioadei Ultimul Maxim glaciar (LGM), care a durat aproximativ de la 26.500 la 19.000 de ani în urmă. I2 a apărut probabil în vestul Europei, deși regiunea sa exactă de origine nu poate fi determinată, deoarece europenii din paleolitic erau vânători-culegători nomazi. Cea mai veche probă I2 recuperată din scheletele arheologice este un bărbat de 13.500 de ani din Grotte du Bichon din Elveția asociat cu cultura Aziliană (vezi Jones și colab. (2015)). Linia sa maternă a fost U5b1h. În octombrie 2016, au fost testate 15 probe de ADN Y din mezolitic european. Dintre aceștia, unul a aparținut haplogrupului C1a2 (în Spania), unul lui F (în Germania), doi lui I* (în Franța) și șase lui I2 (Luxemburg și Suedia), inclusiv lui I2a1* (P37.2), I2a1a1a. (L672), I2a1b (M423) și I2c2 (PF3827). Liniile materne (ADNmt) pe care le-au purtat au fost U2e, U4, U5a1, U5a2 și U5b. Cele patru mostre din Rusia au aparținut haplogrupului Y J*, R1a1 (2x) și R1b1a. Acest lucru arată că a existat deja o anumită diversitate între filiațiile europene mezolitice, deși multe dintre aceste filiații (C1a2, F, I*, J*) sunt acum extrem de rare. Haplogrupul I2a1 pare să fi ieșit din perioada neolitică drept marele câștigător din motive care nu sunt încă clare.
Analiza genomilor neoliticului timpuriu din Anatolia de către Mathieson și colab. (2015) arată că I2c (L596) era deja prezent în nord-vestul Turciei cu 8.500 de ani în urmă și a fost asimilat de fermierii neolitici în drumul lor din Semiluna Fertilă către Europa. Diverse efectuate între 2014 și 2016 au dezvăluit că toate filiațiile europene din mezolitic au fost absorbite de valul de fermieri din Neolitic. Cu toate acestea, I2a s-a remarcat ca fiind cea mai comună dintre ele, însumând 16% din cele 69 de probe de ADN-Y din Neolitic testate. Asta îl plasează ca al doilea cel mai frecvent descendență paternă după G2a, descendența originală a fermierilor anatolieni. I2a1 pare să fi prosperat în mod deosebit în cultura Starčevo–Kőrсs–Criş (6000-4500 î.Hr.) din sud-estul Europei, unde este încă una dintre principalele linii masculine de astăzi. I2a1 a apărut și în cultura Printed-Cardium Pottery (5000-1500 î.Hr.) în vestul Mediteranei. Sardinienii moderni și bascii au moștenit cea mai mare parte a genomului lor de la acei fermieri neolitici din cultura ceramicii tipărite cu cardu.
Deși până acum a fost găsit un singur I2a2 în timpul neoliticului timpuriu sau mijlociu (un I2a2a-M223 din Spania), multe dintre ele au apărut în timpul epocii cuprului și bronzului. Eșantioanele din epoca de cupru și bronz constau dintr-un I2a2, două I2a2a și unul I2a2a1 (CTS616) în Spania, unul I2a2a1b1b2 (S12195) în sudul Rusiei (cultura Yamna), un I2a2a1 (CTS9183) și o cultură I2a2a2a1b1b2 (S12195) în Ungaria (Ungaria) ), șase I2a2b (L38) în Germania (culturi Unetice și Urnfield). Probele I2a1 din epoca cuprului includ un I2a1a1 din nordul Italiei (cultura Remedello), un I2a1 din Ungaria (cultura Vatya). I2c2 a fost găsit și în cultura Unetic din Germania.
Haplogrup I2a1 (P37.2)
Haplogrupul I2a1 este de departe cea mai mare ramură a lui I2 și cea mai strâns legată de culturile neolitice din sud-estul, sud-vestul și nord-vestul Europei.

Haplogrup I2a1a1 (M26)
I2a1a (M26, L158, L159.1/S169.1) a fost cunoscut ca I1b2 până în 2005, I1b1b în 2006-7 și I2a1 din 2008 până în 2010. Se găsește în toată Europa de Vest și atinge frecvențele maxime în rândul sardinilor (37,5%) și bascii (5%), două izolate populației. M26 este limitat geografic la Insulele Britanice, Țările de Jos, Franța, vestul Germaniei, Elveția, Sardinia, Sicilia, coasta de vest a Italiei, Iberia și coasta mediteraneană a Magrebului. Singurele M26 negative pentru mutația L160 sunt limitate la Irlanda.
I2a1a-M26 a fost probabil una dintre principalele linii paterne ale culturilor megalitice din vestul Europei în timpul perioadelor neolitice și calcolitice. I2a1a1a (L672) a fost deja găsit în Suedia mezolitică, ceea ce înseamnă că I2a1a a avut o răspândire foarte largă din Iberia până în Scandinavia în perioada mezolitică. Mai târziu, ei ar fi adoptat agricultura prin amestecarea cu noii veniți din Orientul Apropiat.
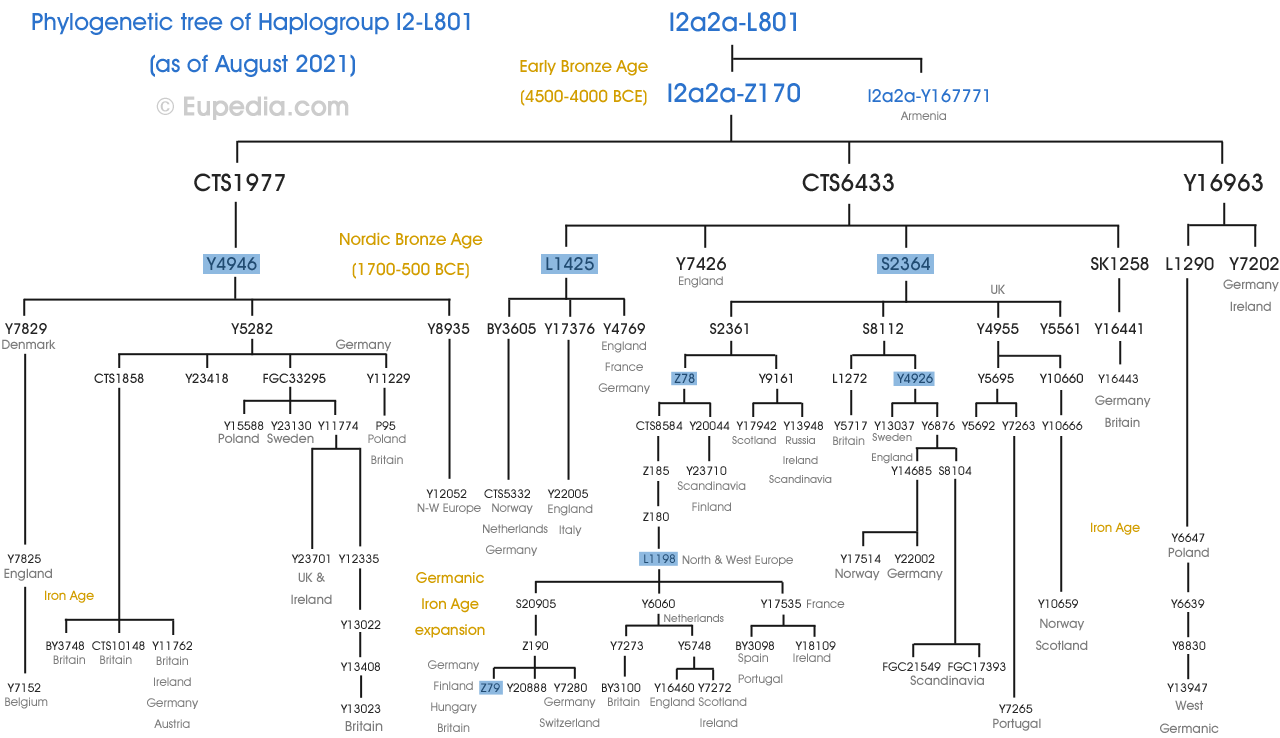
Haplogrup I2a1b (M423)
I2a1b (M423, L178) a fost cunoscut ca I1b până în 2007 și I2a2 din 2008 până în 2010. Subclada principală, reprezentând peste 90% din toate liniile M423 este L621și subclada sa L147.2. Celelalte subclade sunt L41.2(foarte rar) și L161.1(găsește mai ales în Germania și Insulele Britanice).
Haplogrupul I2a1b-L621
Această ramură se găsește în mare parte în țările slave. Frecvențele sale maxime sunt observate în rândul slavilor dinarici (sloveni, croați, bosniaci, sârbi, muntenegreni și macedoneni), precum și în Bulgaria, România, Moldova, vestul Ucrainei și Belarus. De asemenea, este comună într-o măsură mai mică în Albania, Grecia, Ungaria, Slovacia, Polonia și sud-vestul Rusiei. I2-L621 (L147.2+) este cunoscut și ca I2a-Din(pentru Dinaric).
Concentrația mare de I2a1b-L621 în nord-estul României, Moldova și centrul Ucrainei amintește de răspândirea maximă a culturii Cucuteni-Trypillian (4800-3000 î.Hr.). Nicio probă de ADN-Y din această cultură nu a fost testată până în prezent, dar pe măsură ce a evoluat ca o ramificație a culturii Starčevo–Kőrсs–Criș, este probabil ca I2a să fi fost una dintre principalele sale linii paterne, iar un efect fondator ar fi putut crește. frecvența acestuia Cultura Cucuteni-Trypillian a fost cea mai avansată cultură neolitică din Europa înainte de invaziile indo-europene din epoca bronzului și pare să fi avut contacte intense cu cultura stepei înainte de extinderea Yamnei în Balcani și Europa Centrală (vezi istoriile R1a). și R1b). Din anul 3500 î.Hr., la începutul perioadei Yamna în stepa ponto-caspică, poporul Cucuteni-Trypillian a început să se extindă spre est în stepa a ceea ce este acum vestul Ucrainei, părăsind orașele lor (cele mai mari din lume la acea vreme), și adoptând un stil de viață din ce în ce mai nomad ca vecinii lor Yamna. Se poate imagina cu ușurință că oamenii Cucuteni-Trypillian au fost asimilați de vecinii Yamna și că s-au răspândit ca o descendență minoritară alături de haplogrupurile R1a și R1b pe măsură ce au avansat spre Marea Baltică odată cu expansiunea Corded Ware. Alternativ, filiațiile I2-L621 ar fi putut trăi într-o relativă izolare de societatea proto-indo-europeană principală undeva în jurul Ucrainei, Poloniei sau Belarusului, apoi, pe măsură ce secolele și mileniile au trecut, s-ar fi amestecat cu populațiile predominant R1a din jurul lor. Amalgamul rezultat ar fi devenit strămoșii protoslavilor.
În zilele noastre, I2a1 este de cinci până la zece ori mai comun decât G2a în Europa de Sud-Est, în timp ce în perioada neolitică G2a a fost de aproximativ patru ori mai comun. Ce poate explica această inversare completă? La un moment dat în istorie, filiațiile I2a1 par să fi beneficiat de a fi de partea câștigătoare. În afară de un impuls minor de la (ipotetic) alăturarea expansiunii către vest a lui Yamna în Europa, principalul eveniment determinant care a permis I2a1b-L621 să devină o descendență majoră est-europeană au fost probabil migrațiile slave din secolul al VI-lea până în secolul al IX-lea d.Hr aparținând lui I2a1b se încadrează în subcladele L147.2 (aka CTS10228, CTS2180 sau Y3111), despre care se crede că a apărut acum 5.600 de ani (chiar înainte de perioada Yamna și expansiunea Trypillian în stepă), dar are un TMRCA de numai 2.300 ani conform lui Yfull. Minoritatea indivizilor I2a1b-L621 negativi pentru L147.2 se găsesc toți în estul Poloniei, Belarus și vestul Ucrainei, ceea ce sugerează că aici a supraviețuit de la subcladele I2a1b-L147.2 foarte rapid de acum 1900 de ani, ceea ce este în concordanță cu momentul etnogenezei slave, având în vedere că durează câteva secole până când un bărbat poate avea destui descendenți masculini pentru a începe să aibă un impact la scara unei populații. Acest strămoș I2-L147.2 ar avea un asemenea impact asupra populației slave timpurii în creștere, încă mică acum 2.300 de ani, dar în plină expansiune.
După ce triburile germanice care trăiau în estul Germaniei și în Polonia, precum goții, vandalii și burgunzii, au invadat Imperiul Roman, slavii care trăiau mai la est au umplut vidul. După prăbușirea Imperiului Roman de Apus în 476, slavii s-au mutat în Alpii dinarici și în Balcani. Până în secolul al IX-lea, slavii au ocupat toate teritoriile moderne de limbă slavă, cu excepția Balcanilor de Est, sub controlul bulgarilor vorbitori de turcă.
În prezent, țările slave de nord au între 9% (Polonia, Cehia) și 21% (Ucraina) din I2a-L621, în timp ce slavii din sud au între 20% (Bulgaria) și 50% (Bosnia). Procentul mai mare de I2a-Din în sud se datorează, probabil, doar unui alt efect fondator, datorită faptului că slavii de sud au provenit din vestul Ucrainei, unde raportul dintre I2a și R1a a fost mai mare. Practic, tot Dinaric I2a se încadrează în ramura L147.2, iar majoritatea ramificației S17250, care descind dintr-un strămoș patriliniar comun care a trăit cu doar 1.800 de ani în urmă.
Haplogrupul I2a1b-L161.1
Cunoscut în mod obișnuit în cercurile de genealogie genetică ca I2-M423-Insule, L161.1 se găsește la cele mai înalte frecvențe în vestul Irlandei (5-10%) și Highlands scoțiani (1-5%), dar se găsește și la frecvențe joase (> 1%) în toată Europa Centrală și de Vest, din Letonia, Lituania și Belarus până la Insulele Britanice și din Scandinavia până în nord-vestul Spaniei. A fost găsit și în Albania, nordul Greciei, Bulgaria și România.
Cei mai bătrâni indivizi I2a1b-L161.1 cunoscuți sunt bărbatul Loschbour în vârstă de 8.000 de ani din Luxemburgul Mezolitic și un bărbat de 7.800 de ani din Motala, în sudul Suediei. I2a1b-L161.1 a fost probabil împrăștiat în cea mai mare parte a Europei în timpul perioadelor glaciare târzie și postglaciare imediate și, după toate probabilitățile, a integrat societatea neolitică ca toate celelalte linii mezolitice. I2a1b-L161.1 a fost foarte probabil una dintre principalele linii neolitice din Insulele Britanice în timpul perioadei megalitice și, prin urmare, ar fi fost printre oamenii care au construit Stonehenge, Knowth, Newgrange și alte mari situri megalitice. Frecvența sa scăzută astăzi și limitarea sa în marginile de nord-vest și de sud-est ale Europei este, fără îndoială, rezultatul numeroaselor valuri de migrații indo-europene din ultimii 5.000 de ani. Se știe acum că haplogrupul proto-celtic R1b-L21 a ajuns în Irlanda în jurul anului 2000 î.Hr. (vezi Cassidy et al. 2015), la doar câteva secole după ce R1b a sosit pentru prima dată în Europa Centrală. Tranziția a fost rapidă, triburile R1b copleșind populația neolitică din Insulele Britanice în doar câteva secole și împingându-le mai spre vest și spre nord, unde L161.1 supraviețuiește astăzi.
Haplogrupurile I2-L880, I2-L1286 și I2-L1294
Aceste subclade minore se găsesc la frecvențe joase în principal în nord-vestul Europei. La fel ca I2a2 (vezi mai jos), aceste filiații sunt probabil descendenții vânătorilor-culegători din centrul și nord-vestul Europei.
L880 este o subcladă foarte rară identificată în Germania, Luxemburg și Franța.
L1286 a apărut în perioada paleoliticului târziu. Cu toate acestea, 99% dintre transportatorii moderni aparțin subcladei L233 și au un strămoș patriliniar comun cu mai puțin de 2.000 de ani în urmă. L233 este cel mai frecvent în Insulele Britanice, dar este întâlnit ocazional în Țările de Jos, Scandinavia, Germania, Franța și Polonia.
L1294 este, de asemenea, foarte rar și a fost găsit doar în Germania, Franța și Marea Britanie.
Haplogrup I2a2 (P214)
I2a2 (S33/M436/P214, P216/S30, P217/S23, P218/S32, L35/S150, L37/S153, L181) a fost cunoscut ca I1c până în 2005 și I2b până în 2010. Este asociat cu pre-celto-germanică oameni din nord-vestul Europei, cum ar fi constructori de megaliti(5000-1200 î.Hr.). Vârsta sa a fost estimată între 21.000 și 13.000 de ani, ceea ce corespunde perioadei epipaleolitice.
I2a2 se găsește în cea mai mare parte a Europei și pare să fi avut o distribuție pe întreg continentul înainte de sosirea fermierilor din Neolitic. Mai multe probe I2a2 mezolitice au fost identificate până în prezent, mai ales de Mathieson și colab. (2017) Aceasta include indivizi din sudul Germaniei (M223 din circa 7200 î.Hr.), Porțile de Fier dintre Serbia și România (Z161 din cca. 6200 î.Hr.), Letonia (CTS10057 din circa 5500 î.Hr.) și sud-estul Ucrainei (L699 din circa 5400 î.Hr.) și L701 din cca 5200 î.Hr.).
În plus, subclada I2c1 (L1251) a fost identificat recent. Se găsește mai ales în Germania și Anglia și la periferia acestora (Irlanda, Norvegia, Franța, Italia, Polonia). Se crede că această subcladă are aproximativ 6000 de ani, ceea ce o plasează în perioada neoliticului târziu. Poate fi asociat cu vânători-culegători din Germania care au adoptat agricultura după ce au intrat în contact cu fermierii din Orientul Apropiat din cultura ceramicii liniare.
Indivizi celebri
Ramura I2a1b-L621 (slavă)
Ramura I2a2a-Z161 (germanică)
Andrew Johnson(1808-1875), al 17-lea președinte și al 16-lea vicepreședinte al Statelor Unite a fost identificat ca membru al haplogrupului I2a2a (fostul I2b1) pe baza rezultatelor din






